LA CÉLULA
Una célula (del latín cellula, diminutivo de cella, ‘hueco’) es la unidad morfológica y funcional de todo ser vivo. De hecho, la célula es el elemento de menor tamaño que puede considerarse vivo.De este modo, puede clasificarse a los organismos vivos según el número de células que posean: si solo tienen una, se les denomina unicelulares (como pueden ser los protozoos o las bacterias, organismos microscópicos); si poseen más, se les llama pluricelulares. En estos últimos el número de células es variable: de unos pocos cientos, como en algunos nematodos, a cientos de billones (10), como en el caso del ser humano. Las células suelen poseer un tamaño de 10 µm y una masa de 1 ng, si bien existen células mucho mayores.
La teoría celular, propuesta en 1838 para los vegetales y en 1839 para los animales por Matthias Jakob Schleiden y Theodor Schwann, postula que todos los organismos están compuestos por células, y que todas las células derivan de otras precedentes. De este modo, todas las funciones vitales emanan de la maquinaria celular y de la interacción entre células adyacentes; además, la tenencia de la información genética, base de la herencia, en su ADN permite la transmisión de aquella de generación en generación.

Historia y teoría celular
La historia de la biología celular ha estado ligada al desarrollo tecnológico que pudiera sustentar su estudio. De este modo, el primer acercamiento a su morfología se inicia con la popularización del microscopio rudimentario de lentes compuestas en el siglo XVII, se suplementa con diversas técnicas histológicas para microscopía óptica en los siglos XIX y XX y alcanza un mayor nivel resolutivo mediante los estudios de microscopía electrónica, de fluorescencia y confocal, entre otros, ya en el siglo XX. El desarrollo de herramientas moleculares, basadas en el manejo de ácidos nucleicos y enzimas permitieron un análisis más exhaustivo a lo largo del siglo XX.

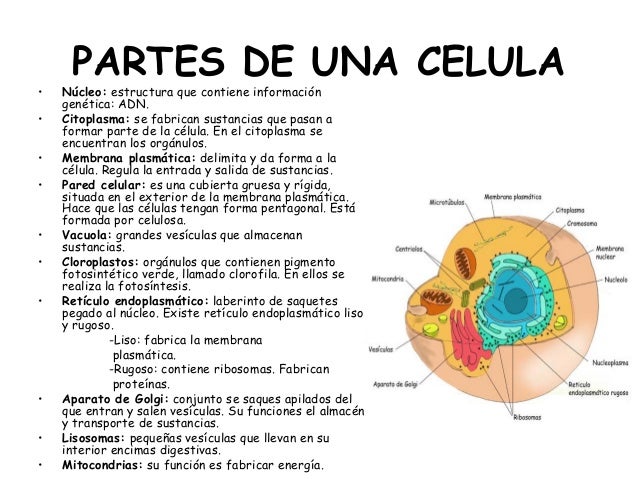
Descubrimiento
Las primeras aproximaciones al estudio de la célula surgieron en el siglo XVII; tras el desarrollo a finales del siglo XVI de los primeros microscopios. Estos permitieron realizar numerosas observaciones, que condujeron en apenas doscientos años a un conocimiento morfológico relativamente aceptable. A continuación se enumera una breve cronología de tales descubrimientos:
1665: Robert Hooke publicó los resultados de sus observaciones sobre tejidos vegetales, como el corcho, realizadas con un microscopio de 50 aumentos construido por él mismo. Este investigador fue el primero que, al ver en esos tejidos unidades que se repetían a modo de celdillas de un panal, las bautizó como elementos de repetición, «células» (del latín cellulae, celdillas). Pero Hooke sólo pudo observar células muertas por lo que no pudo describir las estructuras de su interior.
Década de 1670: Anton van Leeuwenhoek, observó diversas células eucariotas (como protozoos y espermatozoides) y procariotas (bacterias).
1745: John Needham describió la presencia de «animálculos» o «infusorios»; se trataba de organismos unicelulares. Dibujo de la estructura del corcho observado por Robert Hooke bajo su microscopio y tal como aparece publicado en Micrographia.
Década de 1830: Theodor Schwann estudió la célula animal; junto con Matthias Schleiden postularon que las células son las unidades elementales en la formación de las plantas y animales, y que son la base fundamental del proceso vital.
1831: Robert Brown describió el núcleo celular.
1839: Purkinje observó el citoplasma celular.
1850: Rudolf Virchow postuló que todas las células provienen de otras células.
1857: Kölliker identificó las mitocondrias.
1860: Pasteur realizó multitud de estudios sobre el metabolismo de levaduras y sobre la asepsia.
1880: August Weismann descubrió que las células actuales comparten similitud estructural y molecular con células de tiempos remotos.
1931: Ernst Ruska construyó el primer microscopio electrónico de transmisión en la Universidad de Berlín. Cuatro años más tarde, obtuvo un poder de resolución doble a la del microscopio óptico.
1981: Lynn Margulis publica su hipótesis sobre la endosimbiosis serial, que explica el origen de la célula eucariota.


Tipos de células
- 1. Células procariotas
- 2. Células eucariotas
Células procariotas:
Las células procariotas son pequeñas y menos complejas que las eucariotas. Contienen ribosomas pero carecen de sistemas de endomembranas (esto es, orgánulos delimitados por membranas biológicas, como puede ser el núcleo celular). Por ello poseen el material genético en el citosol. Sin embargo, existen excepciones: algunas bacterias fotosintéticas poseen sistemas de membranas internos. También en el Filo Planctomycetes existen organismos como Pirellula que rodean su material genético mediante una membrana intracitoplasmática y Gemmata obscuriglobus que lo rodea con doble membrana. Esta última posee además otros compartimentos internos de membrana, posiblemente conectados con la membrana externa del nucleoide y con la membrana nuclear, que no posee peptidoglucano.
Por lo general podría decirse que los procariotas carecen de citoesqueleto. Sin embargo se ha observado que algunas bacterias, como Bacillus subtilis, poseen proteínas tales como MreB y mbl que actúan de un modo similar a la actina y son importantes en la morfología celular. De gran diversidad, los procariotas sustentan un metabolismo extraordinariamente complejo, en algunos casos exclusivos de ciertos taxa, como algunos grupos de bacterias, lo que incide en su versatilidad ecológica. Los procariotas se clasifican, según Carl Woese, en arqueas y bacterias.
Arqueas

Bacterias

Células eucariotas
La célula eucariota posee compartimentos internos delimitados por membranas. Entre éstos se encuentra el núcleo, delimitado por una doble unidad de membrana, en cuyo interior se encuentra el material genético o ADN que contiene la información necesaria para que la célula pueda llevar a cabo todas las tareas para su supervivencia y reproducción. Entre el núcleo y la membrana plasmática se encuentra el citosol, un gel acuoso que contiene numerosas moléculas que intervienen en funciones estructurales, metabólicas, en la homeostasis, en la señalización. Cabe destacar los ribosomas para la producción de proteínas, el citoesqueleto para la organización interna de la célula y para su movilidad, numerosos enzimas y cofactores para el metabolismo y muchas otras moléculas más. En el citosol también se encuentran los orgánulos, que son compartimentos rodeados por membrana que llevan a cabo funciones como la digestión, respiración, fotosíntesis, metabolismo, transporte intracelular, secreción, producción de energía, almacenamiento, etcétera. Las mitocondrias, los cloroplastos, los peroxisomas, los lisosomas, el retículo endoplasmático, o las vacuolas, entre otros, son orgánulos. El citoplasma es el citosol más los orgánulos que contiene.
Estas se encuentras de manera tradicional dividida en dos tipos células animal y vegetal.

http://www.bionova.org.es/animbio/anim/cellinteract.swf
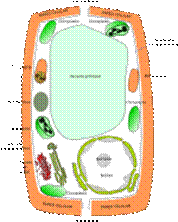
Esquema de los principales componentes de una célula vegetal.
http://cienciasnaturales.es/CELULAVEGETAL.swf

Diagrama de una célula animal, a la izquierda (1. Nucléolo, 2. Núcleo, 3. Ribosoma, 4. Vesícula, 5. Retículo endoplasmático rugoso, 6. Aparato de Golgi, 7. Citoesqueleto (microtúbulos), 8. Retículo endoplasmático liso, 9. Mitocondria, 10. Vacuola, 11. Citoplasma, 12. Lisosoma. 13.Centríolos.).
 | ||||||||||||||||||||||||||||||||||||||||||
Estructura de la célula Membrana Celular
No solo es el límite entre la célula o su entorno sino que también media al contacto y al intercambio entre el citoplasma de una célula y su medio circundante a través de estructuras de transporte. Las membranas celulares son estructuras muy importantes para las células, ya que funcionan como una barrera entre los componentes de la célula y el medio ambiente exterior. La membrana celular no sólo es responsable de crear un muro entre el interior y exterior de la célula, sino que también debe actuar como un umbral a través del cual las moléculas de selección pueden entrar y salir de la celuda cuando sea necesario. La membrana celular es lo que define a la célula y mantiene sus componentes por separado a partir de células u organismos fuera. La membrana celular está compuesta por una doble capa de células de grasa llamado una bicapa lipídica en la que las proteínas de membrana están integradas. La estructura de la bicapa lipídica impide el libre paso de la mayoría de las moléculas dentro y fuera de la célula.

Clasificaciones de proteínas de membrana Las proteínas se suele dividir en las clasificaciones más pequeña de las proteínas integrales, proteínas periféricas, proteínas y lípidos de ruedas.


 
  
Características de algunas moléculas de la superfamilia de las inmunoglobulinas
La extravasación del linfocito en el ganglio linfático ocurre selectivamente al nivel de las vénulas poscapilares endoteliales altas (HEV) presentes en las mucosas del tejido linfoide (igualmente ocurre en las placas de Peyer en el intestino). El desarrollo de estas es una consecuencia de la activación de los linfocitos T y la producción de citocinas como el interferón g. La principal característica de esta extravasación, es la mayor adhesividad del linfocito circulante al endotelio alto durante varios segundos, a diferencia del aplanado, al que solo se asocian durante la fracción de un segundo. La baja afinidad de adhesión hace que por la fuerza del flujo sanguíneo, los linfocitos se separen del endotelio, aunque una pequeña parte pueden adherirse firmemente y a través de las células endoteliales penetren al estroma del ganglio linfático. Esta firme adhesión es posible por la interacción de la L-selectina expresada en los linfocitos T vírgenes con la molécula CD34, proteoglicano sobre las células endoteliales, y con la molécula MadCAM-1 expresada en la mucosa intestinal. Si las células T no interaccionan con el antígeno específico en los ganglios linfáticos, estas salen por los vasos linfáticos eferentes, continúan su recirculación linfática y regresan a la sangre a través del conducto torácico. En el intestino, el tiempo de unión de las células T es más prolongado, lo que provoca que el 50 % de estas penetren en la mucosa.
La inflamación es una compleja serie de reacciones homeostáticas que involucra a los mecanismos inmunológicos humorales y celulares para proteger al organismo. Si esta reacción resulta exagerada o crónica, no cumple su función, y ocurren cambios patológicos.
Se caracteriza por una reacción vascular inicial a un estímulo localizado (reconocimiento antigénico) con liberación de mediadores vasoactivos, una reacción celular de reclutamiento de células inflamatorias (leucocitos inmunocompetentes) que depende de la adhesión leucocitaria, y una reacción tisular en la que los leucocitos liberan mediadores inflamatorios, provocando los efectos deseados (eliminación del antígeno) o no (destrucción tisular).
   | ||||||||||||||||||||||||||||||||||||||||||
"EL CONOCIMIENTO Y LA CIENCIA, PALABRAS QUE DESCRIBEN LA VERDADERA CARA DEL ALMA HUMANA; PORQUE EN ELLAS ESTA LA VERDAD" La critique anonyme